Bacillus and Paenibacillus
Biosynthetic Gene Clusters
Many Bacillus and Paenibacillus
species produce a structurally diverse set of secondary
metabolites; many of these compounds inhibit the growth of
root-associated pathogens. As such, these bacteria and the
compounds they generate hold exceptional promise for protection
of globally essential agricultural products.
Secondary
metabolites are small organic compounds produced by cells that
are not strictly essential for their moment-to-moment survival
but often impart novel properties that enable an organism to
occupy a particular ecological niche. Invariably, the series of
enzymes/proteins necessary for producing these metabolites are
encoded in biosynthetic gene clusters (BGCs). Different types of
BGCs give rise to different classes of secondary metabolites,
including polyketides (PKs), non-ribosomal peptides (NRPs),
ribosomally-synthesized post-translationally modified peptides
(RiPPs), and others. Within classes there is substantial
variation, often depending on the specific identity and
arrangement of modules which compose a given BGC.
Very often, the compounds generated from BGCs have antibiotic
properties against bacteria, fungi, and/or oomycetes. In
collaboration with the Liles
and Noel
laboratories, we are evaluating the antibiosis properties (breadth
of organisms inhibited, potency of inhibition, etc.) of many
hundreds of Bacillus and Paenibacillus strains
spanning multiple species. From complete genome sequence data, we
are able to identify the BGCs encoded within each strain. We
extract metabolites from activecultures, evaluate their
antibiotic properties, and use metabolomic tools (e.g., LC-MS/MS)
to identify and characterize these compounds. Strains from B.
velezensis are particularly prolific in the production of
secondary metabolites, and the compounds that many of them
generate give rise to broad spectrum abilities to inhibit
notorious agricultural pathogens, including Phytophthora
nicotianae, Globisporangium ultimum, Rhizoctonia
solani, Fusarium oxysporum, etc.
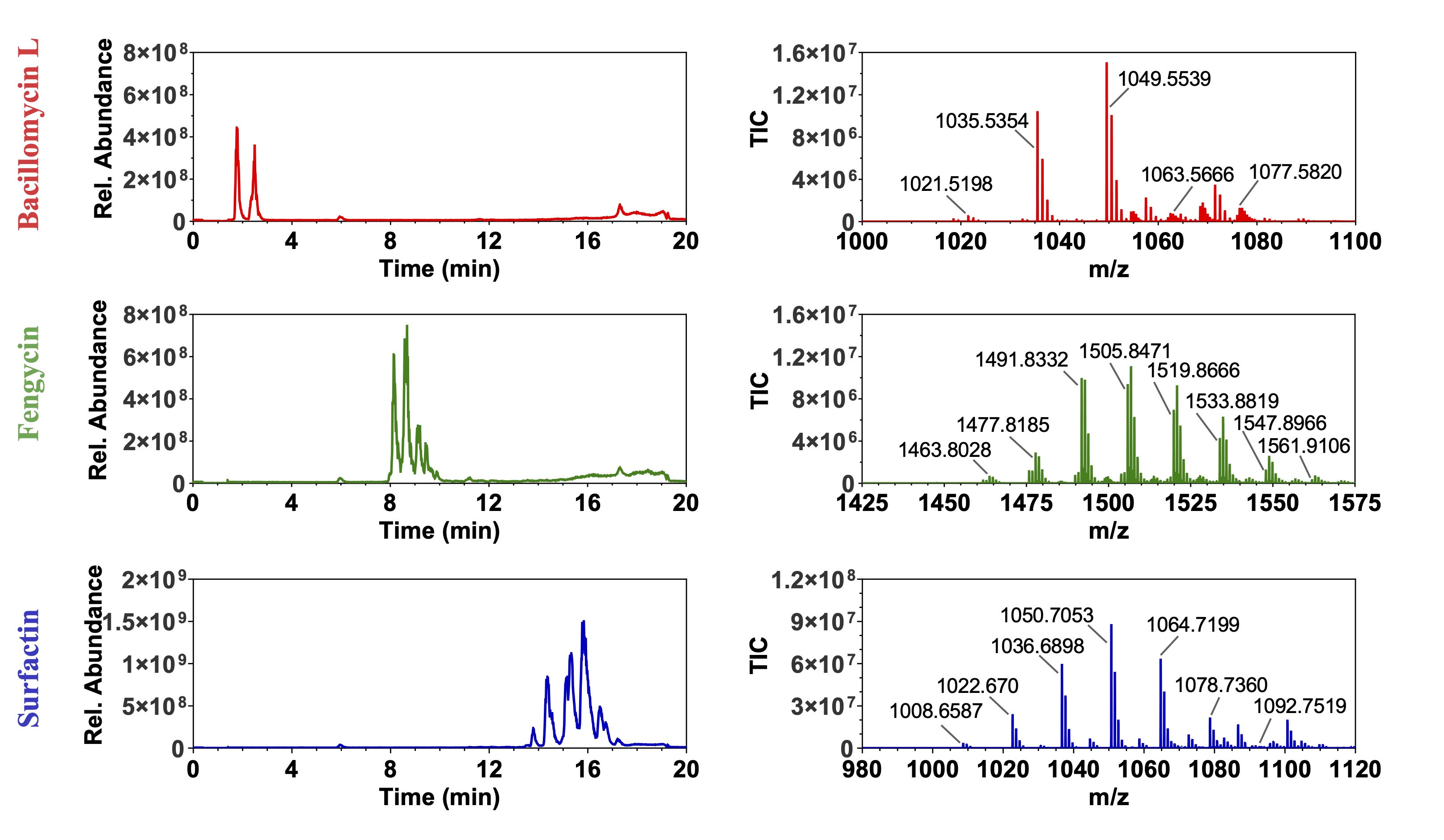
Image: LC (left) and MS (right) for three
lipopeptides from B. velezensis JJ334.
Multiple variants of each are present; they appear in m/z
increments of 14 amu.
This is due to variation in fatty acid chain length and core
peptide substitution (e.g., Val v. Ile).